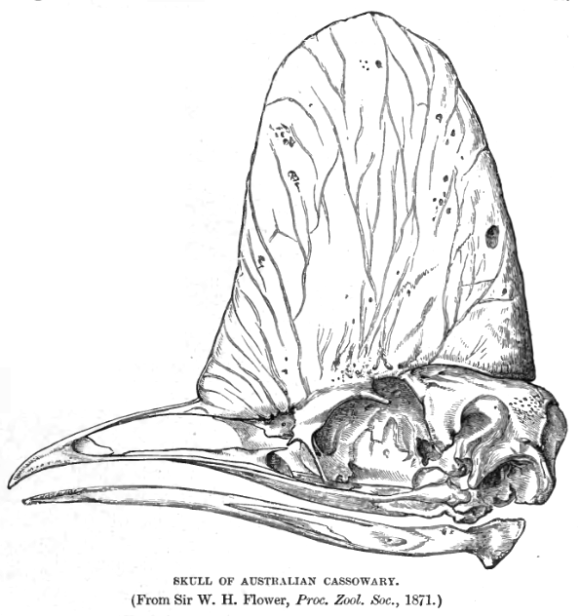
Nuevos artículos 12/03/2019 1 comentario en Los casuarios regulan la temperatura corporal con el casco Los casuarios regulan la temperatura corporal con el casco 05/03/2019 1 comentario en La nariz que parece una mano y hace la tarea de los ojos La nariz que parece una mano y hace la tarea de los ojos 26/02/2019 1 comentario en La hibernación limita el tamaño encefálico La hibernación limita el tamaño encefálico 21/12/2018 1 comentario en Vivir a toda prisa, estivar durante meses Vivir a toda prisa, estivar durante meses 14/02/2018 12 comentarios en Los colibríes no se envenenan con agua Los colibríes no se envenenan con agua 08/02/2018 8 comentarios en El pájaro más pequeño que existe El pájaro más pequeño que existe 27/09/2017 3 comentarios en La salamandra solar La salamandra solar 13/09/2017 No hay comentarios en Aves que migran Aves que migran 06/09/2017 2 comentarios en Gran buceador, volador mediocre Gran buceador, volador mediocre 30/08/2017 3 comentarios en El cuervo acuático El cuervo acuático 23/08/2017 1 comentario en El precio de la migración El precio de la migración 16/08/2017 1 comentario en Serpientes Serpientes 1 2 3 … 17 ›